





Abstract
Land snails of the Papua New Guinean camaenid genus Rhynchotrochus are described on the basis of shell shape, shell colour and banding pattern. We compared morphological and anatomical differentiation in three of the species, viz. R. albocarinatus, R. taylorianus and R. williamsi. The three species can be distinguished by their shell colour and banding pattern, as well as by their shell and reproductive morphology. Rhynchotrochus albocarinatus has a white, subglobose shell with up to five bands and a white to chocolate-brown outer lip. The penis is longer than the vas deferens and the length of the oviduct (LO) is approximately one-third that of the vagina. The subdepressed, trochiform shell of R. taylorianus is highly variable with respect to shell colour and banding pattern. The penis is approximately as long as the epiphallus and shorter than the vas deferens; the LO is approximately two-thirds that of the vagina. Rhynchotrochus williamsi has a white, unbanded, globose shell with a white to light pink outer lip and a dark protoconch. The penis is markedly shorter than the vas deferens or epiphallus which are approximately of the same length; the oviduct is approximately half as long as the vagina. Within R. taylorianus, there is substantial variation in shell colour, shell banding pattern and reproductive morphology that may confuse identification. We also document on one specimen of R. albocarinatus that has two interconnected penises. Since both penises were not connected to the spermoviduct, copulation as a male should have been prevented in this specimen.
INTRODUCTION
Snails show remarkable interspecific variations in shell colouration, banding pattern and ornamentation (e.g. Schilthuizen, 2003), which can be used for taxonomic purposes. Nevertheless, numerous studies have documented high intraspecific, geographical variation in the form and colouration of shells, and that shell types are often associated with particular habitats (references in Goodfriend, 1986). Such variation may limit the use of shell characters as taxonomic markers. Similarly, several studies (e.g. Davis, Chen & Yu, 1994; Wilke, Pfenninger & Davis, 2002) have demonstrated unusually high intraspecific variability in anatomical characters, indicating poor taxonomic resolution of these characters, although qualitative characters of genitalia are extensively used for taxonomic and systematic studies (see also Fiorentino, Manganelli & Giusti, 2008). The combined analysis of shell and genital characters may reveal characters that are reliable taxonomic markers whereas other characters show low taxonomic resolution.
The Camaenidae (as defined by Zilch, 1959–1960) are a helicoid family of land snails with a disjunct distribution. Seven genera are found in America where they are distributed from Costa Rica, the Greater and Lesser Antilles to northern Argentina, while 28 genera occur in Japan, southern China, the Philippines, SE Asia, New Guinea and Australia (Wurtz, 1955; Solem, 1992a). Because of a lack of characters unique to the family, the Camaenidae are a taxonomically confused group and literature dealing with the intrageneric systematics and phylogeny is limited. A cladistic analysis suggested that the Camaenidae are a monophyletic family with an oval genital orifice and the absence of a penial sheath as synapomorphies (Cuezzo, 2003). However, Scott (1996), Wade, Mordan & Clarke (2001), Wade, Mordan & Naggs (2006) and Wade et al. (2007) suggested that the Camaenidae is a polyphyletic taxon, with the American taxa closely related to the Helicidae and Helminthoglyptidae (Scott, 1996) or separated into two groups (Wade et al., 2007), while the Asian and Australasian taxa appear to form a complex with the Bradybaenidae (Wade et al., 2007). Within the Camaenidae, especially the American (e.g. Wurtz, 1955; Bishop, 1979; Roth, 1988) and Australian (e.g. Solem, 1979, 1981a, b, 1984, 1985, 1992a, b, 1993, 1997) genera have received some attention from taxonomists but Southeast Asian taxa have largely been ignored.
Land snails of the genus Rhynchotrochus Möllendorff, 1895 are characterized by a depressed, trochoid and usually keeled shell with a downward-depressed aperture (Clench & Turner, 1966). The outer lip has a ‘papuinoid notch’, i.e. a dent-like structure named after the subfamily Panuininae to which the genus belongs (Clench & Turner, 1966). Rhynchotrochus are arboricole species that are mostly confined to Papua New Guinea and its satellite islands. Two species reach the Bismarck Archipelago and one occurs in northern Queensland (Australia). Compared to several other helicoid taxa, the genital system of Rhynchotrochus is very simple and lacks mucous glands, a dart-sac and a bursa stalk diverticulum. The epiphallus has a short appendix and there is a single spermatheca. The anatomy is known for only four of the 20 Rhynchotrochus species, viz. R. taylorianus (Adams & Reeve, 1850), R. kubaryi (Möllendorff, 1895), R. wiegmanni (v. Martens, 1894) and R. macgillivrayi (Forbes, 1852) (Clench & Turner, 1966) and, with the exception of R. taylorianus, only one or two preserved specimens are available. Because the anatomy is unknown for most of the Rhynchotrochus species and the genital morphology of the four above mentioned species is very similar (Clench & Turner, 1966), genital characters have never been evaluated as taxonomic markers in this genus. Hence, the taxonomy and systematics of Rhynchotrochus is based solely on characters of the shell, namely shape, colour and banding pattern.
In this study, we used multivariate statistics on shell and genital characters to evaluate their potential as taxonomic markers in three Rhynchotrochus species from Papua New Guinea, viz. R. albocarinatus (Smith, 1887), R. williamsi (Clench & Archer, 1936) and R. taylorianus (Adams & Reeve, 1850), and to evaluate intraspecific variation in shell and genital morphology in R. taylorianus. We also report on an individual of R. albocarinatus that shows an anomaly in the male reproductive organs.
MATERIAL AND METHODS
Three-hundred Rhynchotrochus specimens were collected from nine localities in Papua New Guinea and stored in 70% ethanol (Table 1). Specimens were preliminarily identified by their shell shape, shell colour and shell-banding pattern following Clench & Turner (1966) (Fig. 1). The soft body was removed from the shell. We measured shell height (Sh), shell width (Sw), aperture height (Ah) and aperture width (Aw) to the nearest 0.01 mm using callipers (Fig. 2). For R. taylorianus, we also measured the width of the aperture notch (An). We scored the colour of the body whorl, the remaining whorls, the protoconch and of the lip, and counted the number of bands on the body whorl. From the proximal reproductive organs the length of the penis (LPe), epiphallus (LEp), vas deferens (LVd), vagina (LVa), oviduct (LO) and the bursa plus pedunculus of the bursa (LPed) (hereafter referred to as the bursa copulatrix) were measured to the nearest 0.01 mm with callipers (Fig. 3).

Frontal (left) and top view (right) of the shells of three Rhynchotrochus species. A.R. albocarinatus from Laing Island. B.R. williamsi from Laing Island. C.R. taylorianus from Madang. D.R. taylorianus from Bam Island. E.R. taylorianus from Boisa Island 4.
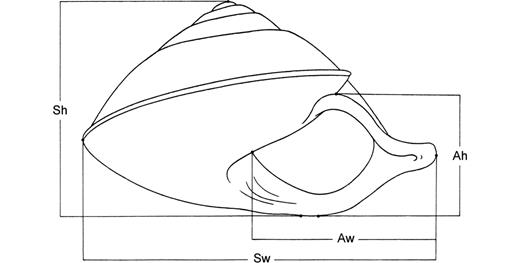
Camera lucida drawing of the frontal view of a shell of Rhynchotrochus albocarinatus with indication of the measured characters. Abbreviations: Ah, aperture height; Aw, aperture width; Sh, shell height; Sw, shell width.
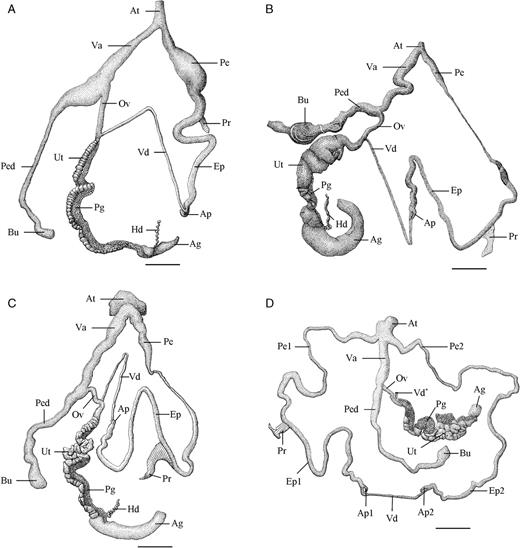
Camera lucida drawings of the proximal reproductive organs of three Rhynchotrochus species. A.R. williamsi from Laing Island. B.R. taylorianus from Laing Island. C.R. albocarinatus from Boisa Island 1. D. R. albocarinatus specimen with two sets of male proximal genitalia. Scale bar = 5 mm. Abbreviations: Ag, albumen gland; Ap, epiphallus appendix; At, genital atrium; Bu, bursa of the bursa copulatrix; Ep, epiphallus; Hd, hermaphroditic duct; Ov, oviduct; Pe, penis; Ped, pedunculus of the bursa copulatrix; Pg, prostate gland; Pr, penis retractor muscle; Ut, uterus; Va, vagina; Vd, vas deferens.
Sampling localities, collection date, shell ground colour, shell banding pattern and number of specimens collected (N) for three Rhynchotrochus species from Papua New Guinea.
| Species | Collection date | N | Shell ground colour | Shell banding pattern | Lip colour | Protoconch colour |
|---|---|---|---|---|---|---|
| Rhynchotrochus albocarinatus | ||||||
| Laing Island (Madang prov.) | 21/05/1976 | 80 | Ivory white | 0–5 narrow bands | Dark brown | Light yellow to brownish |
| Rhynchotrochus williamsi | ||||||
| Laing Island (Madang prov.) | 21/05/1976 | 4 | White | No bands | Pink | Dark brown to black |
| Rhynchotrochus taylorianus | ||||||
| Boisa Island 1 (Madang prov.) | 24/05/1977 | 35 | Yellowish | No bands | White | Dark purple |
| Boisa Island 2 (Madang prov.) | 26/05/1977 | 32 | Yellowish | No bands | White | Dark purple |
| Boisa Island 3 (Madang prov.) | 7–8/06/1978 | 35 | Yellowish | No bands | White | Dark purple |
| Boisa Island 4 (Madang prov.) | 22/06/1977 | 23 | Yellowish | No bands | White | Dark purple |
| Bam Island (East Sepik prov.) | 14/06/1978 | 65 | Creamy white | 2–8 narrow bands | White | White |
| Blup Blup (East Sepik prov.) | ??/06/1978 | 19 | White | 3 broad bands | White | White |
| Madang Province (unspecified) | 1976 | 3 | Purplish | 4 bands | Dark purple to black | Dark purple |
| Tarawa | 1976 | 4 | White to yellowish | 3 broad bands | White | White |
| Species | Collection date | N | Shell ground colour | Shell banding pattern | Lip colour | Protoconch colour |
|---|---|---|---|---|---|---|
| Rhynchotrochus albocarinatus | ||||||
| Laing Island (Madang prov.) | 21/05/1976 | 80 | Ivory white | 0–5 narrow bands | Dark brown | Light yellow to brownish |
| Rhynchotrochus williamsi | ||||||
| Laing Island (Madang prov.) | 21/05/1976 | 4 | White | No bands | Pink | Dark brown to black |
| Rhynchotrochus taylorianus | ||||||
| Boisa Island 1 (Madang prov.) | 24/05/1977 | 35 | Yellowish | No bands | White | Dark purple |
| Boisa Island 2 (Madang prov.) | 26/05/1977 | 32 | Yellowish | No bands | White | Dark purple |
| Boisa Island 3 (Madang prov.) | 7–8/06/1978 | 35 | Yellowish | No bands | White | Dark purple |
| Boisa Island 4 (Madang prov.) | 22/06/1977 | 23 | Yellowish | No bands | White | Dark purple |
| Bam Island (East Sepik prov.) | 14/06/1978 | 65 | Creamy white | 2–8 narrow bands | White | White |
| Blup Blup (East Sepik prov.) | ??/06/1978 | 19 | White | 3 broad bands | White | White |
| Madang Province (unspecified) | 1976 | 3 | Purplish | 4 bands | Dark purple to black | Dark purple |
| Tarawa | 1976 | 4 | White to yellowish | 3 broad bands | White | White |
Sampling localities, collection date, shell ground colour, shell banding pattern and number of specimens collected (N) for three Rhynchotrochus species from Papua New Guinea.
| Species | Collection date | N | Shell ground colour | Shell banding pattern | Lip colour | Protoconch colour |
|---|---|---|---|---|---|---|
| Rhynchotrochus albocarinatus | ||||||
| Laing Island (Madang prov.) | 21/05/1976 | 80 | Ivory white | 0–5 narrow bands | Dark brown | Light yellow to brownish |
| Rhynchotrochus williamsi | ||||||
| Laing Island (Madang prov.) | 21/05/1976 | 4 | White | No bands | Pink | Dark brown to black |
| Rhynchotrochus taylorianus | ||||||
| Boisa Island 1 (Madang prov.) | 24/05/1977 | 35 | Yellowish | No bands | White | Dark purple |
| Boisa Island 2 (Madang prov.) | 26/05/1977 | 32 | Yellowish | No bands | White | Dark purple |
| Boisa Island 3 (Madang prov.) | 7–8/06/1978 | 35 | Yellowish | No bands | White | Dark purple |
| Boisa Island 4 (Madang prov.) | 22/06/1977 | 23 | Yellowish | No bands | White | Dark purple |
| Bam Island (East Sepik prov.) | 14/06/1978 | 65 | Creamy white | 2–8 narrow bands | White | White |
| Blup Blup (East Sepik prov.) | ??/06/1978 | 19 | White | 3 broad bands | White | White |
| Madang Province (unspecified) | 1976 | 3 | Purplish | 4 bands | Dark purple to black | Dark purple |
| Tarawa | 1976 | 4 | White to yellowish | 3 broad bands | White | White |
| Species | Collection date | N | Shell ground colour | Shell banding pattern | Lip colour | Protoconch colour |
|---|---|---|---|---|---|---|
| Rhynchotrochus albocarinatus | ||||||
| Laing Island (Madang prov.) | 21/05/1976 | 80 | Ivory white | 0–5 narrow bands | Dark brown | Light yellow to brownish |
| Rhynchotrochus williamsi | ||||||
| Laing Island (Madang prov.) | 21/05/1976 | 4 | White | No bands | Pink | Dark brown to black |
| Rhynchotrochus taylorianus | ||||||
| Boisa Island 1 (Madang prov.) | 24/05/1977 | 35 | Yellowish | No bands | White | Dark purple |
| Boisa Island 2 (Madang prov.) | 26/05/1977 | 32 | Yellowish | No bands | White | Dark purple |
| Boisa Island 3 (Madang prov.) | 7–8/06/1978 | 35 | Yellowish | No bands | White | Dark purple |
| Boisa Island 4 (Madang prov.) | 22/06/1977 | 23 | Yellowish | No bands | White | Dark purple |
| Bam Island (East Sepik prov.) | 14/06/1978 | 65 | Creamy white | 2–8 narrow bands | White | White |
| Blup Blup (East Sepik prov.) | ??/06/1978 | 19 | White | 3 broad bands | White | White |
| Madang Province (unspecified) | 1976 | 3 | Purplish | 4 bands | Dark purple to black | Dark purple |
| Tarawa | 1976 | 4 | White to yellowish | 3 broad bands | White | White |
All data were log10-transformed to approximate normality. To reduce allometric effects, a multivariate analysis of covariance (MANCOVA) with Sh as covariate was used to analyse morphometrical differences in shell and genitalia among the three species. Differences in Sh were analysed by one-way analysis of variance (ANOVA). Post hoc comparisons were made by Scheffé tests. A multivariate ANOVA yielded virtually the same results and will not be further discussed. A canonical discriminant analysis (CDA) was used to extract discriminant functions by which an individual could be assigned to one of the taxa of the analysis. Group differences were evaluated by computing the proportion of (in)correctly classified individuals in each group. For R. taylorianus, we collected individuals from eight localities (Table 1). From six localities we collected sufficient individuals to evaluate morphometrical differences among localities using MANCOVA and CDA.
Because we could not measure the genitalia of all individuals, the dataset was analysed in three separate ways: (1) one in which only the shell characters were considered; (2) one including the genital characters only and (3) one in which the shell and genital characters were analysed together. All analyses were performed using Statistica v.5.5 (StatSoft, Inc., Tulsa). Detailed data lists are available upon request.
RESULTS
Basic statistics of the shell and genital characters are given in Table 2. Overall, we observed more variation in the genital than in the shell characters. A summary of the morphological differences and similarities between the three species is given in Table 3.
Mean ± SE (in mm) of the morphological variables of the shell and the reproductive organs in Rhynchotrochus albocarinatus, R. williamsi and R. taylorianus from Papua New Guinea.
| Species | Shell | Reproductive organs | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SH | SW | AH | AW | AN | LPE | LEP | LVD | LVA | LPED | LO | |
| R. albocarinatus | 16.8 ± 0.15 | 22.2 ± 0.15 | 11.9 ± 0.1 | 11.4 ± 0.1 | – | 34.5 ± 1.1 | 27.4 ± 0.7 | 23.4 ± 0.4 | 15.4 ± 0.3 | 21.0 ± 0.4 | 4.8 ± 0.2 |
| R. williamsi | 19.8 ± 0.3 | 25.5 ± 0.4 | 13.6 ± 0.2 | 13.0 ± 0.3 | – | 17.1 ± 0.9 | 26.0 ± 1.9 | 28.8 ± 1.5 | 11.1 ± 0.9 | 29.5 ± 2.5 | 6.9 ± 0.4 |
| R. taylorianus (overall) | 17.1 ± 0.1 | 25.4 ± 0.2 | 12.1 ± 0.1 | 13.1 ± 0.1 | 8.9 ± 0.1 | 17.4 ± 0.2 | 19.2 ± 0.2 | 23.6 ± 0.3 | 9.8 ± 0.1 | 25.5 ± 0.4 | 6.1 ± 0.1 |
| Boisa Island 1 | 15.9 ± 0.1 | 24.2 ± 0.3 | 11.4 ± 0.1 | 12.7 ± 0.3 | 8.1 ± 0.1 | 17.3 ± 0.2 | 18.4 ± 0.5 | 21.0 ± 0.4 | 8.9 ± 0.2 | 23.2 ± 0.6 | 5.7 ± 0.2 |
| Boisa Island 2 | 16.0 ± 0.1 | 24.2 ± 0.2 | 11.5 ± 0.1 | 12.3 ± 0.1 | 8.4 ± 0.1 | 17.2 ± 0.2 | 17.6 ± 0.4 | 22.2 ± 0.5 | 9.6 ± 0.4 | 23.9 ± 0.8 | 6.0 ± 0.3 |
| Boisa Island 3 | 16.1 ± 0.2 | 24.6 ± 0.4 | 11.5 ± 0.1 | 12.6 ± 0.1 | 8.3 ± 0.3 | 17.6 ± 0.3 | 18.4 ± 0.4 | 21.7 ± 0.4 | 9.7 ± 0.2 | 25.0 ± 0.7 | 5.5 ± 0.2 |
| Boisa Island 4 | 15.6 ± 0.2 | 23.2 ± 0.2 | 11.0 ± 0.1 | 11.8 ± 0.1 | 8.0 ± 0.1 | 17.7 ± 0.3 | 19.4 ± 0.8 | 22.2 ± 1.1 | 9.5 ± 0.4 | 22.3 ± 2.1 | 6.6 ± 0.6 |
| Bam Island | 18.3 ± 0.1 | 26.0 ± 0.2 | 12.5 ± 0.1 | 13.4 ± 0.1 | 9.5 ± 0.1 | 17.0 ± 1.0 | 21.0 ± 0.4 | 26.2 ± 0.4 | 10.5 ± 0.2 | 29.4 ± 0.6 | 6.5 ± 0.2 |
| Blup Blup | 20.2 ± 0.4 | 30.7 ± 0.6 | 15.2 ± 0.2 | 16.1 ± 0.3 | 10.9 ± 0.2 | 16.2 ± 1.3 | 19.8 ± 0.8 | 30.0 ± 1.3 | 11.9 ± 0.8 | 30.4 ± 1.8 | 7.7 ± 0.4 |
| Madang | 17.6 ± 0.7 | 28.6 ± 0.5 | 13.1 ± 0.3 | 14.8 ± 0.3 | 9.2 ± 0.2 | 15.0 ± 0.9 | 16.1 ± 3.9 | 27.6 ± 1.2 | 10.7 ± 2.0 | 22.1 | 5.5 ± 0.8 |
| Tarawa | 17.4 ± 1.2 | 28.2 ± 1.1 | 13.4 ± 0.5 | 13.9 ± 0.2 | 10.5 ± 0.5 | 15.0 ± 0.9 | 20.8 ± 0.5 | 23.1 ± 1.8 | 7.1 ± 0.1 | 21.7 | 6.5 ± 0.1 |
| Species | Shell | Reproductive organs | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SH | SW | AH | AW | AN | LPE | LEP | LVD | LVA | LPED | LO | |
| R. albocarinatus | 16.8 ± 0.15 | 22.2 ± 0.15 | 11.9 ± 0.1 | 11.4 ± 0.1 | – | 34.5 ± 1.1 | 27.4 ± 0.7 | 23.4 ± 0.4 | 15.4 ± 0.3 | 21.0 ± 0.4 | 4.8 ± 0.2 |
| R. williamsi | 19.8 ± 0.3 | 25.5 ± 0.4 | 13.6 ± 0.2 | 13.0 ± 0.3 | – | 17.1 ± 0.9 | 26.0 ± 1.9 | 28.8 ± 1.5 | 11.1 ± 0.9 | 29.5 ± 2.5 | 6.9 ± 0.4 |
| R. taylorianus (overall) | 17.1 ± 0.1 | 25.4 ± 0.2 | 12.1 ± 0.1 | 13.1 ± 0.1 | 8.9 ± 0.1 | 17.4 ± 0.2 | 19.2 ± 0.2 | 23.6 ± 0.3 | 9.8 ± 0.1 | 25.5 ± 0.4 | 6.1 ± 0.1 |
| Boisa Island 1 | 15.9 ± 0.1 | 24.2 ± 0.3 | 11.4 ± 0.1 | 12.7 ± 0.3 | 8.1 ± 0.1 | 17.3 ± 0.2 | 18.4 ± 0.5 | 21.0 ± 0.4 | 8.9 ± 0.2 | 23.2 ± 0.6 | 5.7 ± 0.2 |
| Boisa Island 2 | 16.0 ± 0.1 | 24.2 ± 0.2 | 11.5 ± 0.1 | 12.3 ± 0.1 | 8.4 ± 0.1 | 17.2 ± 0.2 | 17.6 ± 0.4 | 22.2 ± 0.5 | 9.6 ± 0.4 | 23.9 ± 0.8 | 6.0 ± 0.3 |
| Boisa Island 3 | 16.1 ± 0.2 | 24.6 ± 0.4 | 11.5 ± 0.1 | 12.6 ± 0.1 | 8.3 ± 0.3 | 17.6 ± 0.3 | 18.4 ± 0.4 | 21.7 ± 0.4 | 9.7 ± 0.2 | 25.0 ± 0.7 | 5.5 ± 0.2 |
| Boisa Island 4 | 15.6 ± 0.2 | 23.2 ± 0.2 | 11.0 ± 0.1 | 11.8 ± 0.1 | 8.0 ± 0.1 | 17.7 ± 0.3 | 19.4 ± 0.8 | 22.2 ± 1.1 | 9.5 ± 0.4 | 22.3 ± 2.1 | 6.6 ± 0.6 |
| Bam Island | 18.3 ± 0.1 | 26.0 ± 0.2 | 12.5 ± 0.1 | 13.4 ± 0.1 | 9.5 ± 0.1 | 17.0 ± 1.0 | 21.0 ± 0.4 | 26.2 ± 0.4 | 10.5 ± 0.2 | 29.4 ± 0.6 | 6.5 ± 0.2 |
| Blup Blup | 20.2 ± 0.4 | 30.7 ± 0.6 | 15.2 ± 0.2 | 16.1 ± 0.3 | 10.9 ± 0.2 | 16.2 ± 1.3 | 19.8 ± 0.8 | 30.0 ± 1.3 | 11.9 ± 0.8 | 30.4 ± 1.8 | 7.7 ± 0.4 |
| Madang | 17.6 ± 0.7 | 28.6 ± 0.5 | 13.1 ± 0.3 | 14.8 ± 0.3 | 9.2 ± 0.2 | 15.0 ± 0.9 | 16.1 ± 3.9 | 27.6 ± 1.2 | 10.7 ± 2.0 | 22.1 | 5.5 ± 0.8 |
| Tarawa | 17.4 ± 1.2 | 28.2 ± 1.1 | 13.4 ± 0.5 | 13.9 ± 0.2 | 10.5 ± 0.5 | 15.0 ± 0.9 | 20.8 ± 0.5 | 23.1 ± 1.8 | 7.1 ± 0.1 | 21.7 | 6.5 ± 0.1 |
Mean ± SE (in mm) of the morphological variables of the shell and the reproductive organs in Rhynchotrochus albocarinatus, R. williamsi and R. taylorianus from Papua New Guinea.
| Species | Shell | Reproductive organs | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SH | SW | AH | AW | AN | LPE | LEP | LVD | LVA | LPED | LO | |
| R. albocarinatus | 16.8 ± 0.15 | 22.2 ± 0.15 | 11.9 ± 0.1 | 11.4 ± 0.1 | – | 34.5 ± 1.1 | 27.4 ± 0.7 | 23.4 ± 0.4 | 15.4 ± 0.3 | 21.0 ± 0.4 | 4.8 ± 0.2 |
| R. williamsi | 19.8 ± 0.3 | 25.5 ± 0.4 | 13.6 ± 0.2 | 13.0 ± 0.3 | – | 17.1 ± 0.9 | 26.0 ± 1.9 | 28.8 ± 1.5 | 11.1 ± 0.9 | 29.5 ± 2.5 | 6.9 ± 0.4 |
| R. taylorianus (overall) | 17.1 ± 0.1 | 25.4 ± 0.2 | 12.1 ± 0.1 | 13.1 ± 0.1 | 8.9 ± 0.1 | 17.4 ± 0.2 | 19.2 ± 0.2 | 23.6 ± 0.3 | 9.8 ± 0.1 | 25.5 ± 0.4 | 6.1 ± 0.1 |
| Boisa Island 1 | 15.9 ± 0.1 | 24.2 ± 0.3 | 11.4 ± 0.1 | 12.7 ± 0.3 | 8.1 ± 0.1 | 17.3 ± 0.2 | 18.4 ± 0.5 | 21.0 ± 0.4 | 8.9 ± 0.2 | 23.2 ± 0.6 | 5.7 ± 0.2 |
| Boisa Island 2 | 16.0 ± 0.1 | 24.2 ± 0.2 | 11.5 ± 0.1 | 12.3 ± 0.1 | 8.4 ± 0.1 | 17.2 ± 0.2 | 17.6 ± 0.4 | 22.2 ± 0.5 | 9.6 ± 0.4 | 23.9 ± 0.8 | 6.0 ± 0.3 |
| Boisa Island 3 | 16.1 ± 0.2 | 24.6 ± 0.4 | 11.5 ± 0.1 | 12.6 ± 0.1 | 8.3 ± 0.3 | 17.6 ± 0.3 | 18.4 ± 0.4 | 21.7 ± 0.4 | 9.7 ± 0.2 | 25.0 ± 0.7 | 5.5 ± 0.2 |
| Boisa Island 4 | 15.6 ± 0.2 | 23.2 ± 0.2 | 11.0 ± 0.1 | 11.8 ± 0.1 | 8.0 ± 0.1 | 17.7 ± 0.3 | 19.4 ± 0.8 | 22.2 ± 1.1 | 9.5 ± 0.4 | 22.3 ± 2.1 | 6.6 ± 0.6 |
| Bam Island | 18.3 ± 0.1 | 26.0 ± 0.2 | 12.5 ± 0.1 | 13.4 ± 0.1 | 9.5 ± 0.1 | 17.0 ± 1.0 | 21.0 ± 0.4 | 26.2 ± 0.4 | 10.5 ± 0.2 | 29.4 ± 0.6 | 6.5 ± 0.2 |
| Blup Blup | 20.2 ± 0.4 | 30.7 ± 0.6 | 15.2 ± 0.2 | 16.1 ± 0.3 | 10.9 ± 0.2 | 16.2 ± 1.3 | 19.8 ± 0.8 | 30.0 ± 1.3 | 11.9 ± 0.8 | 30.4 ± 1.8 | 7.7 ± 0.4 |
| Madang | 17.6 ± 0.7 | 28.6 ± 0.5 | 13.1 ± 0.3 | 14.8 ± 0.3 | 9.2 ± 0.2 | 15.0 ± 0.9 | 16.1 ± 3.9 | 27.6 ± 1.2 | 10.7 ± 2.0 | 22.1 | 5.5 ± 0.8 |
| Tarawa | 17.4 ± 1.2 | 28.2 ± 1.1 | 13.4 ± 0.5 | 13.9 ± 0.2 | 10.5 ± 0.5 | 15.0 ± 0.9 | 20.8 ± 0.5 | 23.1 ± 1.8 | 7.1 ± 0.1 | 21.7 | 6.5 ± 0.1 |
| Species | Shell | Reproductive organs | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SH | SW | AH | AW | AN | LPE | LEP | LVD | LVA | LPED | LO | |
| R. albocarinatus | 16.8 ± 0.15 | 22.2 ± 0.15 | 11.9 ± 0.1 | 11.4 ± 0.1 | – | 34.5 ± 1.1 | 27.4 ± 0.7 | 23.4 ± 0.4 | 15.4 ± 0.3 | 21.0 ± 0.4 | 4.8 ± 0.2 |
| R. williamsi | 19.8 ± 0.3 | 25.5 ± 0.4 | 13.6 ± 0.2 | 13.0 ± 0.3 | – | 17.1 ± 0.9 | 26.0 ± 1.9 | 28.8 ± 1.5 | 11.1 ± 0.9 | 29.5 ± 2.5 | 6.9 ± 0.4 |
| R. taylorianus (overall) | 17.1 ± 0.1 | 25.4 ± 0.2 | 12.1 ± 0.1 | 13.1 ± 0.1 | 8.9 ± 0.1 | 17.4 ± 0.2 | 19.2 ± 0.2 | 23.6 ± 0.3 | 9.8 ± 0.1 | 25.5 ± 0.4 | 6.1 ± 0.1 |
| Boisa Island 1 | 15.9 ± 0.1 | 24.2 ± 0.3 | 11.4 ± 0.1 | 12.7 ± 0.3 | 8.1 ± 0.1 | 17.3 ± 0.2 | 18.4 ± 0.5 | 21.0 ± 0.4 | 8.9 ± 0.2 | 23.2 ± 0.6 | 5.7 ± 0.2 |
| Boisa Island 2 | 16.0 ± 0.1 | 24.2 ± 0.2 | 11.5 ± 0.1 | 12.3 ± 0.1 | 8.4 ± 0.1 | 17.2 ± 0.2 | 17.6 ± 0.4 | 22.2 ± 0.5 | 9.6 ± 0.4 | 23.9 ± 0.8 | 6.0 ± 0.3 |
| Boisa Island 3 | 16.1 ± 0.2 | 24.6 ± 0.4 | 11.5 ± 0.1 | 12.6 ± 0.1 | 8.3 ± 0.3 | 17.6 ± 0.3 | 18.4 ± 0.4 | 21.7 ± 0.4 | 9.7 ± 0.2 | 25.0 ± 0.7 | 5.5 ± 0.2 |
| Boisa Island 4 | 15.6 ± 0.2 | 23.2 ± 0.2 | 11.0 ± 0.1 | 11.8 ± 0.1 | 8.0 ± 0.1 | 17.7 ± 0.3 | 19.4 ± 0.8 | 22.2 ± 1.1 | 9.5 ± 0.4 | 22.3 ± 2.1 | 6.6 ± 0.6 |
| Bam Island | 18.3 ± 0.1 | 26.0 ± 0.2 | 12.5 ± 0.1 | 13.4 ± 0.1 | 9.5 ± 0.1 | 17.0 ± 1.0 | 21.0 ± 0.4 | 26.2 ± 0.4 | 10.5 ± 0.2 | 29.4 ± 0.6 | 6.5 ± 0.2 |
| Blup Blup | 20.2 ± 0.4 | 30.7 ± 0.6 | 15.2 ± 0.2 | 16.1 ± 0.3 | 10.9 ± 0.2 | 16.2 ± 1.3 | 19.8 ± 0.8 | 30.0 ± 1.3 | 11.9 ± 0.8 | 30.4 ± 1.8 | 7.7 ± 0.4 |
| Madang | 17.6 ± 0.7 | 28.6 ± 0.5 | 13.1 ± 0.3 | 14.8 ± 0.3 | 9.2 ± 0.2 | 15.0 ± 0.9 | 16.1 ± 3.9 | 27.6 ± 1.2 | 10.7 ± 2.0 | 22.1 | 5.5 ± 0.8 |
| Tarawa | 17.4 ± 1.2 | 28.2 ± 1.1 | 13.4 ± 0.5 | 13.9 ± 0.2 | 10.5 ± 0.5 | 15.0 ± 0.9 | 20.8 ± 0.5 | 23.1 ± 1.8 | 7.1 ± 0.1 | 21.7 | 6.5 ± 0.1 |
Summarizing table for the morphological differences in shell and reproductive organs in Rhynchotrochus albocarinatus, R. williamsi and R. taylorianus from Papua New Guinea.
| Species | R. albocarinatus | R. williamsi | R. taylorianus |
|---|---|---|---|
| Shell colour | |||
| Ground colour | Ivory white | White | White/yellow/purple |
| Banding pattern | 0–5 bands | Unbanded | 0–8 bands |
| Lip colour | Dark brown | Pink | White/purple/black |
| Protoconch colour | Yellow/brown | Brown/black | White/purple |
| Shell shape | |||
| Shell height | = | > | = |
| Shell width | < | = | = |
| Aperture height | = | = | = |
| Aperture width | < | = | = |
| Reproductive organs | |||
| Penis length | > | = | = |
| Epiphallus length | = | = | < |
| Vas deferens length | = | > | = |
| Vagina length | > | = | = |
| Oviduct length | < | = | = |
| Pedunculus + bursa length | < | = | = |
| Species | R. albocarinatus | R. williamsi | R. taylorianus |
|---|---|---|---|
| Shell colour | |||
| Ground colour | Ivory white | White | White/yellow/purple |
| Banding pattern | 0–5 bands | Unbanded | 0–8 bands |
| Lip colour | Dark brown | Pink | White/purple/black |
| Protoconch colour | Yellow/brown | Brown/black | White/purple |
| Shell shape | |||
| Shell height | = | > | = |
| Shell width | < | = | = |
| Aperture height | = | = | = |
| Aperture width | < | = | = |
| Reproductive organs | |||
| Penis length | > | = | = |
| Epiphallus length | = | = | < |
| Vas deferens length | = | > | = |
| Vagina length | > | = | = |
| Oviduct length | < | = | = |
| Pedunculus + bursa length | < | = | = |
‘>’indicates that the character was significantly higher, wider or longer than in both other species, ‘<’indicates that the character was significantly lower, slender or shorter than in both other species, ‘ = ’indicates no statistical significant differences. Diagnostic characters (i.e. characters that show no overlap) are given in bold.
Summarizing table for the morphological differences in shell and reproductive organs in Rhynchotrochus albocarinatus, R. williamsi and R. taylorianus from Papua New Guinea.
| Species | R. albocarinatus | R. williamsi | R. taylorianus |
|---|---|---|---|
| Shell colour | |||
| Ground colour | Ivory white | White | White/yellow/purple |
| Banding pattern | 0–5 bands | Unbanded | 0–8 bands |
| Lip colour | Dark brown | Pink | White/purple/black |
| Protoconch colour | Yellow/brown | Brown/black | White/purple |
| Shell shape | |||
| Shell height | = | > | = |
| Shell width | < | = | = |
| Aperture height | = | = | = |
| Aperture width | < | = | = |
| Reproductive organs | |||
| Penis length | > | = | = |
| Epiphallus length | = | = | < |
| Vas deferens length | = | > | = |
| Vagina length | > | = | = |
| Oviduct length | < | = | = |
| Pedunculus + bursa length | < | = | = |
| Species | R. albocarinatus | R. williamsi | R. taylorianus |
|---|---|---|---|
| Shell colour | |||
| Ground colour | Ivory white | White | White/yellow/purple |
| Banding pattern | 0–5 bands | Unbanded | 0–8 bands |
| Lip colour | Dark brown | Pink | White/purple/black |
| Protoconch colour | Yellow/brown | Brown/black | White/purple |
| Shell shape | |||
| Shell height | = | > | = |
| Shell width | < | = | = |
| Aperture height | = | = | = |
| Aperture width | < | = | = |
| Reproductive organs | |||
| Penis length | > | = | = |
| Epiphallus length | = | = | < |
| Vas deferens length | = | > | = |
| Vagina length | > | = | = |
| Oviduct length | < | = | = |
| Pedunculus + bursa length | < | = | = |
‘>’indicates that the character was significantly higher, wider or longer than in both other species, ‘<’indicates that the character was significantly lower, slender or shorter than in both other species, ‘ = ’indicates no statistical significant differences. Diagnostic characters (i.e. characters that show no overlap) are given in bold.
Species differences: shells
Sh was significantly different among the three species (F = 9.19; df = 2,163; P < 0.001). Shells of R. williamsi were higher than those of both other species (Scheffé test: P < 0.001). The Sh of R. albocarinatus and R. taylorianus was not significantly different. The MANCOVA analysis revealed significant differences in the other shell characters among the three species (Wilks' Lambda = 0.44; df = 6,586; P < 0.001). The shells of R. williamsi and R. taylorianus did not differ in Sw and Aw but R. albocarinatus had a smaller Sw and Aw (Scheffé test: both P < 0.001).
In a CDA of the analysed shell measurements 8% of the shells were classified incorrectly (Table 4; Fig. 4A). Thirteen per cent of the R. albocarinatus shells were classified as R. taylorianus whereas 4% of the R. taylorianus shells were classified as R. albocarinatus. Three of the four R. williamsi shells were classified as R. albocarinatus and the fourth shell was classified as R. taylorianus.
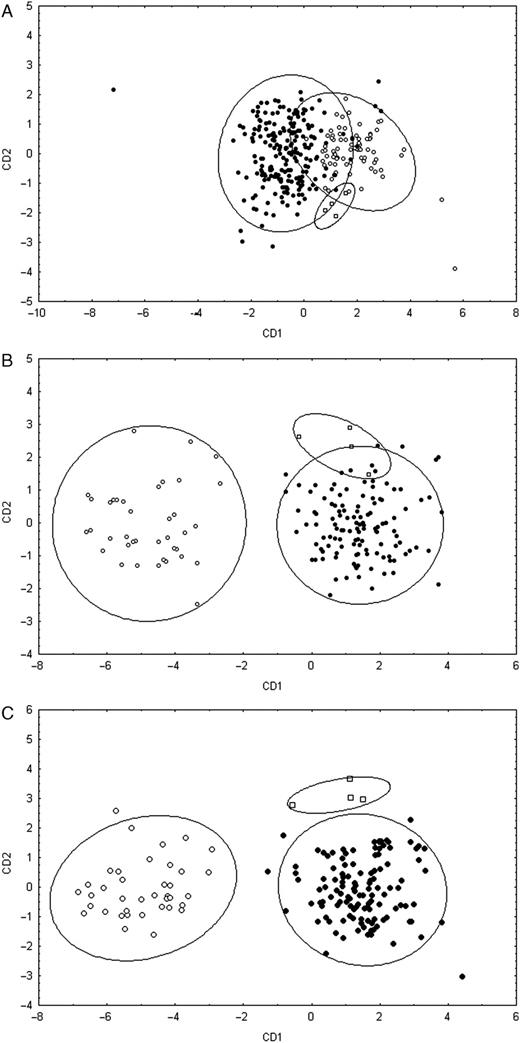
Plot of the first two canonical discriminant functions for the datasets of the shell characters (A), the characters of the reproductive organs (B) and the characters of the shell and reproductive organs combined (C). Symbols: open circles, R. albocarinatus; squares, R. williamsi; filled circles, R. taylorianus. Ellipses represent the 95% confidence intervals.
Classification matrix of the canonical discriminant analyses of the shell characters, the characters of the reproductive organs and the combined characters of shell and reproductive organs in three Rhynchotrochus species from Papua New Guinea.
| Classified as: | Percentage of individuals classified correctly | |||
|---|---|---|---|---|
| R. albocarinatus | R. williamsi | R. taylorianus | ||
| Shell | ||||
| R. albocarinatus | 69 | 0 | 10 | 87 |
| R. williamsi | 3 | 0 | 1 | 0 |
| R. taylorianus | 9 | 0 | 206 | 96 |
| Reproductive organs | ||||
| R. albocarinatus | 38 | 0 | 0 | 100 |
| R. williamsi | 0 | 2 | 2 | 50 |
| R. taylorianus | 0 | 0 | 125 | 100 |
| Shell + reproductive organs | ||||
| R. albocarinatus | 38 | 0 | 0 | 100 |
| R. williamsi | 0 | 4 | 0 | 100 |
| R. taylorianus | 0 | 0 | 124 | 100 |
| Classified as: | Percentage of individuals classified correctly | |||
|---|---|---|---|---|
| R. albocarinatus | R. williamsi | R. taylorianus | ||
| Shell | ||||
| R. albocarinatus | 69 | 0 | 10 | 87 |
| R. williamsi | 3 | 0 | 1 | 0 |
| R. taylorianus | 9 | 0 | 206 | 96 |
| Reproductive organs | ||||
| R. albocarinatus | 38 | 0 | 0 | 100 |
| R. williamsi | 0 | 2 | 2 | 50 |
| R. taylorianus | 0 | 0 | 125 | 100 |
| Shell + reproductive organs | ||||
| R. albocarinatus | 38 | 0 | 0 | 100 |
| R. williamsi | 0 | 4 | 0 | 100 |
| R. taylorianus | 0 | 0 | 124 | 100 |
Classification matrix of the canonical discriminant analyses of the shell characters, the characters of the reproductive organs and the combined characters of shell and reproductive organs in three Rhynchotrochus species from Papua New Guinea.
| Classified as: | Percentage of individuals classified correctly | |||
|---|---|---|---|---|
| R. albocarinatus | R. williamsi | R. taylorianus | ||
| Shell | ||||
| R. albocarinatus | 69 | 0 | 10 | 87 |
| R. williamsi | 3 | 0 | 1 | 0 |
| R. taylorianus | 9 | 0 | 206 | 96 |
| Reproductive organs | ||||
| R. albocarinatus | 38 | 0 | 0 | 100 |
| R. williamsi | 0 | 2 | 2 | 50 |
| R. taylorianus | 0 | 0 | 125 | 100 |
| Shell + reproductive organs | ||||
| R. albocarinatus | 38 | 0 | 0 | 100 |
| R. williamsi | 0 | 4 | 0 | 100 |
| R. taylorianus | 0 | 0 | 124 | 100 |
| Classified as: | Percentage of individuals classified correctly | |||
|---|---|---|---|---|
| R. albocarinatus | R. williamsi | R. taylorianus | ||
| Shell | ||||
| R. albocarinatus | 69 | 0 | 10 | 87 |
| R. williamsi | 3 | 0 | 1 | 0 |
| R. taylorianus | 9 | 0 | 206 | 96 |
| Reproductive organs | ||||
| R. albocarinatus | 38 | 0 | 0 | 100 |
| R. williamsi | 0 | 2 | 2 | 50 |
| R. taylorianus | 0 | 0 | 125 | 100 |
| Shell + reproductive organs | ||||
| R. albocarinatus | 38 | 0 | 0 | 100 |
| R. williamsi | 0 | 4 | 0 | 100 |
| R. taylorianus | 0 | 0 | 124 | 100 |
The shells of R. albocarinatus (Fig. 1A) were subglobose and had a white ground colour with 0–5 narrow bands. The lip was dark brown and the protoconch was light yellow to brownish. The shells of R. williamsi (Fig. 1B) were globose, had a white ground colour and bands were absent or very faint. The lip was pink and the protoconch was dark brown to black. We found five different colour types within R. taylorianus (Table 1; Fig. 1C–E). Individuals of the four localities of Boisa Island were of the same shell colour type, viz. unbanded, yellowish and with a dark purple protoconch. Individuals from the four other localities were characterized by a different shell colour type. Shells of R. taylorianus were trochiform (Fig. 1C–E) and the shell ground colour was white, yellowish or purplish; shells were banded or unbanded; the lip colour was either white or dark purple to black and the protoconch colour was either white or dark purple (Table 1).
Species differences: reproductive organs
The MANCOVA analysis revealed significant morphometrical differences in the genitalia among the three species (Wilks' Lambda = 0.12; df = 12,316; P < 0.001). The penis and vagina were significantly longer, and the bursa copulatrix and the oviduct were significantly shorter, in R. albocarinatus (Fig. 3C) than in both other species that showed no significant differences for these characters. Rhynchotrochus williamsi (Fig. 3A) had a longer vas deferens than both other species which did not differ significantly in this character. Finally, R. taylorianus (Fig. 3B) had a significantly shorter epiphallus than both other species. The LEp was not significantly different between R. albocarinatus and R. williamsi.
Almost all individuals were classified correctly in a CDA of the analysed reproductive organs measurements (Table 4; Fig. 4B). Only two R. williamsi individuals were classified as R. taylorianus based on their genitalia.
Species differences: combined analysis of shell and reproductive organs
The MANCOVA analysis on the combined data of the shell and reproductive organs showed significant differences among the three species (Wilks' Lambda = 0.11; df = 18,308; P < 0.001). The Scheffé tests revealed the same differences among the three species as the analyses on the shell characters or reproductive characters only. In a CDA analysis of the analysed shell and genital measurements all individuals were classified correctly (Table 4; Fig. 4C).
Interpopulation differences in shell and genital morphology in R. taylorianus
The six populations of R. taylorianus differed significantly in both shell morphology (Wilks' Lambda = 0.59; df = 15,552; P < 0.001) and genital morphology (Wilks' Lambda = 0.51; df = 30,446; P < 0.001). Significant differences were also found for the combined dataset (Wilks' Lambda = 0.33; df = 45,481; P < 0.001). For the shell characters, the Scheffé tests revealed that shell morphology of individuals from the four localities of Boisa Island was not significantly different but that these individuals were significantly smaller than individuals from Bam Island and Blup Blup (all P < 0.001). Shell morphology of individuals of the latter two localities was not significantly different.
There were no statistical significant differences among the populations for the LPe, vagina and oviduct. Individuals from Blup Blup had a significantly longer vas deferens and bursa copulatrix than individuals from Boisa Island 1, 2, 3 and 4 and individuals from Bam Island had a longer epiphallus than individuals from Boisa Island 2. All other comparisons were not significant.
In a CDA of the analysed shell or genital measurements many individuals were misclassified (Table 5; Fig. 5). Only a high number of individuals of Bam Island and Blup Blup could be correctly assigned using shell characters or characters of the reproductive organs only (Table 5; Figs. 5A, B). Combining shell and genital characters yielded a very high number of correctly classified individuals for Bam Island and Blup Blup. From the localities Boisa Island 1, 2 and 3, still more than half the number of individuals were incorrectly classified when combining shell and reproductive characters (Table 5; Fig. 5C).
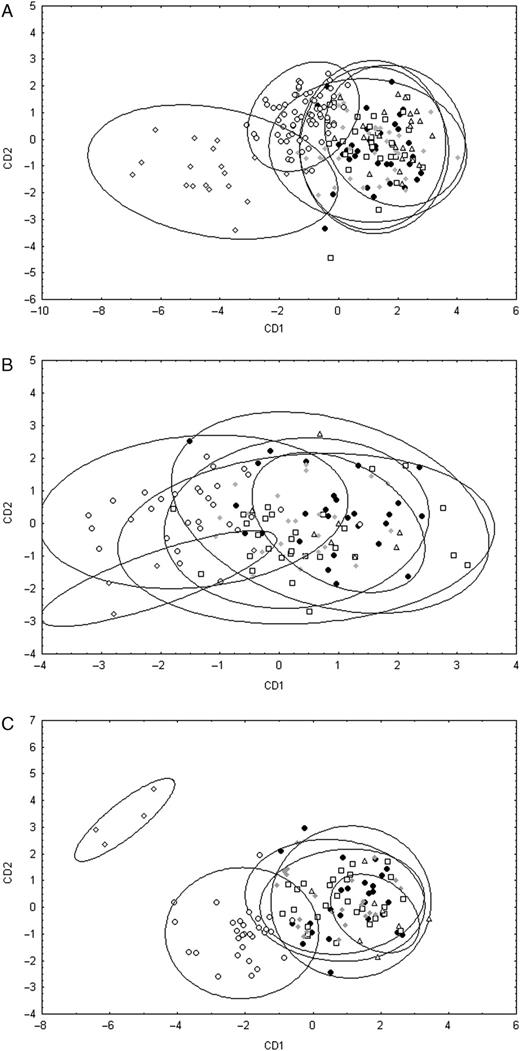
Plot of the first two canonical discriminant functions for the datasets of the shell characters (A), the characters of the reproductive organs (B) and the characters of the shell and reproductive organs combined (C) of the six populations of R. taylorianus. Symbols: filled circles, Boisa Island 1; squares, Boisa Island 2; grey diamonds, Boisa Island 3; triangles, Boisa Island 4; open circles, Bam Island; open diamonds, Blup Blup. Ellipses represent the 95% confidence intervals.
Classification matrix of the canonical discriminant analyses of the shell characters, the characters of the reproductive organs and the combined characters of shell and reproductive organs in six localities of Rhynchotrochus taylorianus from Papua New Guinea.
| Classified as: | Percentage of individuals classified correctly | ||||||
|---|---|---|---|---|---|---|---|
| BI1 | BI2 | BI3 | BI4 | BAI | Blup Blup | ||
| Shell | |||||||
| Boisa Island 1 (BI1) | 15 | 3 | 9 | 6 | 2 | 0 | 43 |
| Boisa Island 2 (BI2) | 11 | 6 | 7 | 4 | 4 | 0 | 19 |
| Boisa Island 3 (BI3) | 13 | 4 | 5 | 3 | 10 | 0 | 14 |
| Boisa Island 4 (BI4) | 5 | 1 | 3 | 12 | 2 | 0 | 52 |
| Bam Island (BAI) | 0 | 0 | 0 | 0 | 65 | 0 | 100 |
| Blup Blup | 0 | 0 | 0 | 0 | 2 | 17 | 89 |
| Reproductive organs | |||||||
| Boisa Island 1 (BI1) | 15 | 8 | 1 | 0 | 3 | 0 | 56 |
| Boisa Island 2 (BI2) | 7 | 14 | 3 | 1 | 4 | 0 | 48 |
| Boisa Island 3 (BI3) | 4 | 7 | 10 | 0 | 4 | 0 | 40 |
| Boisa Island 4 (BI4) | 1 | 1 | 2 | 3 | 1 | 0 | 38 |
| Bam Island (BAI) | 0 | 1 | 3 | 0 | 26 | 0 | 87 |
| Blup Blup | 0 | 1 | 0 | 0 | 1 | 2 | 50 |
| Shell + reproductive organs | |||||||
| Boisa Island 1 (BI1) | 13 | 9 | 2 | 1 | 2 | 0 | 48 |
| Boisa Island 2 (BI2) | 8 | 14 | 6 | 0 | 1 | 0 | 48 |
| Boisa Island 3 (BI3) | 6 | 9 | 8 | 0 | 1 | 0 | 33 |
| Boisa Island 4 (BI4) | 0 | 3 | 2 | 3 | 0 | 0 | 38 |
| Bam Island (BAI) | 0 | 1 | 1 | 0 | 28 | 0 | 93 |
| Blup Blup | 0 | 0 | 0 | 0 | 0 | 4 | 100 |
| Classified as: | Percentage of individuals classified correctly | ||||||
|---|---|---|---|---|---|---|---|
| BI1 | BI2 | BI3 | BI4 | BAI | Blup Blup | ||
| Shell | |||||||
| Boisa Island 1 (BI1) | 15 | 3 | 9 | 6 | 2 | 0 | 43 |
| Boisa Island 2 (BI2) | 11 | 6 | 7 | 4 | 4 | 0 | 19 |
| Boisa Island 3 (BI3) | 13 | 4 | 5 | 3 | 10 | 0 | 14 |
| Boisa Island 4 (BI4) | 5 | 1 | 3 | 12 | 2 | 0 | 52 |
| Bam Island (BAI) | 0 | 0 | 0 | 0 | 65 | 0 | 100 |
| Blup Blup | 0 | 0 | 0 | 0 | 2 | 17 | 89 |
| Reproductive organs | |||||||
| Boisa Island 1 (BI1) | 15 | 8 | 1 | 0 | 3 | 0 | 56 |
| Boisa Island 2 (BI2) | 7 | 14 | 3 | 1 | 4 | 0 | 48 |
| Boisa Island 3 (BI3) | 4 | 7 | 10 | 0 | 4 | 0 | 40 |
| Boisa Island 4 (BI4) | 1 | 1 | 2 | 3 | 1 | 0 | 38 |
| Bam Island (BAI) | 0 | 1 | 3 | 0 | 26 | 0 | 87 |
| Blup Blup | 0 | 1 | 0 | 0 | 1 | 2 | 50 |
| Shell + reproductive organs | |||||||
| Boisa Island 1 (BI1) | 13 | 9 | 2 | 1 | 2 | 0 | 48 |
| Boisa Island 2 (BI2) | 8 | 14 | 6 | 0 | 1 | 0 | 48 |
| Boisa Island 3 (BI3) | 6 | 9 | 8 | 0 | 1 | 0 | 33 |
| Boisa Island 4 (BI4) | 0 | 3 | 2 | 3 | 0 | 0 | 38 |
| Bam Island (BAI) | 0 | 1 | 1 | 0 | 28 | 0 | 93 |
| Blup Blup | 0 | 0 | 0 | 0 | 0 | 4 | 100 |
Classification matrix of the canonical discriminant analyses of the shell characters, the characters of the reproductive organs and the combined characters of shell and reproductive organs in six localities of Rhynchotrochus taylorianus from Papua New Guinea.
| Classified as: | Percentage of individuals classified correctly | ||||||
|---|---|---|---|---|---|---|---|
| BI1 | BI2 | BI3 | BI4 | BAI | Blup Blup | ||
| Shell | |||||||
| Boisa Island 1 (BI1) | 15 | 3 | 9 | 6 | 2 | 0 | 43 |
| Boisa Island 2 (BI2) | 11 | 6 | 7 | 4 | 4 | 0 | 19 |
| Boisa Island 3 (BI3) | 13 | 4 | 5 | 3 | 10 | 0 | 14 |
| Boisa Island 4 (BI4) | 5 | 1 | 3 | 12 | 2 | 0 | 52 |
| Bam Island (BAI) | 0 | 0 | 0 | 0 | 65 | 0 | 100 |
| Blup Blup | 0 | 0 | 0 | 0 | 2 | 17 | 89 |
| Reproductive organs | |||||||
| Boisa Island 1 (BI1) | 15 | 8 | 1 | 0 | 3 | 0 | 56 |
| Boisa Island 2 (BI2) | 7 | 14 | 3 | 1 | 4 | 0 | 48 |
| Boisa Island 3 (BI3) | 4 | 7 | 10 | 0 | 4 | 0 | 40 |
| Boisa Island 4 (BI4) | 1 | 1 | 2 | 3 | 1 | 0 | 38 |
| Bam Island (BAI) | 0 | 1 | 3 | 0 | 26 | 0 | 87 |
| Blup Blup | 0 | 1 | 0 | 0 | 1 | 2 | 50 |
| Shell + reproductive organs | |||||||
| Boisa Island 1 (BI1) | 13 | 9 | 2 | 1 | 2 | 0 | 48 |
| Boisa Island 2 (BI2) | 8 | 14 | 6 | 0 | 1 | 0 | 48 |
| Boisa Island 3 (BI3) | 6 | 9 | 8 | 0 | 1 | 0 | 33 |
| Boisa Island 4 (BI4) | 0 | 3 | 2 | 3 | 0 | 0 | 38 |
| Bam Island (BAI) | 0 | 1 | 1 | 0 | 28 | 0 | 93 |
| Blup Blup | 0 | 0 | 0 | 0 | 0 | 4 | 100 |
| Classified as: | Percentage of individuals classified correctly | ||||||
|---|---|---|---|---|---|---|---|
| BI1 | BI2 | BI3 | BI4 | BAI | Blup Blup | ||
| Shell | |||||||
| Boisa Island 1 (BI1) | 15 | 3 | 9 | 6 | 2 | 0 | 43 |
| Boisa Island 2 (BI2) | 11 | 6 | 7 | 4 | 4 | 0 | 19 |
| Boisa Island 3 (BI3) | 13 | 4 | 5 | 3 | 10 | 0 | 14 |
| Boisa Island 4 (BI4) | 5 | 1 | 3 | 12 | 2 | 0 | 52 |
| Bam Island (BAI) | 0 | 0 | 0 | 0 | 65 | 0 | 100 |
| Blup Blup | 0 | 0 | 0 | 0 | 2 | 17 | 89 |
| Reproductive organs | |||||||
| Boisa Island 1 (BI1) | 15 | 8 | 1 | 0 | 3 | 0 | 56 |
| Boisa Island 2 (BI2) | 7 | 14 | 3 | 1 | 4 | 0 | 48 |
| Boisa Island 3 (BI3) | 4 | 7 | 10 | 0 | 4 | 0 | 40 |
| Boisa Island 4 (BI4) | 1 | 1 | 2 | 3 | 1 | 0 | 38 |
| Bam Island (BAI) | 0 | 1 | 3 | 0 | 26 | 0 | 87 |
| Blup Blup | 0 | 1 | 0 | 0 | 1 | 2 | 50 |
| Shell + reproductive organs | |||||||
| Boisa Island 1 (BI1) | 13 | 9 | 2 | 1 | 2 | 0 | 48 |
| Boisa Island 2 (BI2) | 8 | 14 | 6 | 0 | 1 | 0 | 48 |
| Boisa Island 3 (BI3) | 6 | 9 | 8 | 0 | 1 | 0 | 33 |
| Boisa Island 4 (BI4) | 0 | 3 | 2 | 3 | 0 | 0 | 38 |
| Bam Island (BAI) | 0 | 1 | 1 | 0 | 28 | 0 | 93 |
| Blup Blup | 0 | 0 | 0 | 0 | 0 | 4 | 100 |
Anomaly of the male reproductive organs in R. albocarinatus
One specimen of R. albocarinatus had an abnormal male reproductive tract (Fig. 3D). The specimen had two penises and two epiphalli that were interconnected by a very short vas deferens that was not connected to the spermoviduct. Each of the penises inserted at a side of the genital atrium. A penial retractor was only present at the end of one of the penises. A small blind-ended vas deferens was present at the spermoviduct. The female reproductive organs appeared normally developed.
DISCUSSION
The identification, taxonomy and systematics of the land snails of the genus Rhynchotrochus is complicated by (1) a high intraspecific variation in shell colour and banding pattern in some of the species, (2) a lack of knowledge on the morphology of the reproductive organs for 16 of the 20 currently described species and (3) a seemingly high similarity of the reproductive morphology of the four species for which the reproductive morphology currently is known (Clench & Turner, 1966). Moreover, the development of stylommatophoran reproductive organs is subject to considerable intraspecific, individual, seasonal and physiological variation (Jordaens et al., 2002, and references therein). Also within the Camaenidae, several studies showed much intraspecific variation in the size and shape of the reproductive organs (Tomiyama, 1988b; Marcos, 1992) and of the shell (Tomiyama, 1988a) which may reduce the reliability of shell and genital morphological characters as taxonomic marker. Therefore, Clench & Turner (1966) claimed that the reproductive organs have little taxonomic value within Rhynchotrochus. For instance, Clench & Turner (1966) mention R. albocarinatus as most closely related to R. williamsi and only differing in shell colouration. We showed that both species can also be discriminated by their reproductive morphology.
Thus, our study shows that, even when the shell and reproductive organs show high intraspecific morphological variation, they may provide suitable taxonomic characters to discriminate among Rhynchotrochus species. Whereas 8% of the individuals were incorrectly classified using shell characters only, the reproductive organs allowed a better classification of the species, while the combined dataset of shell and genital characters allowed the correct identification of all individuals. Remarkably, characters of the female reproductive organs (i.e. the LVa, pedunculus of the bursa copulatrix and oviduct) seem especially diagnostic whereas a high overlap was observed for the characters of the male reproductive organs (Tables 2 and 3).
Rhynchotrocus albocarinatus is characterized by a long penis and vagina and a short bursa copulatrix and oviduct. The penis of this species is much longer than the epiphallus or vas deferens, and consists of a short, muscular proximal part and a very long, thin distal part. The vas deferens is slightly shorter than the epiphallus. The LO is approximately one-third that of the vagina. According to Clench & Turner (1966), the shell of R. albocarinatus is subglobose and ivory white with the older whorls a light yellowish brown to pinkish brown. The shell has a series of light to dark-brown spiral bands, both below and above the periphery. The bands may be narrow and more or less evenly spaced, or two or more bands may coalesce (Clench & Turner, 1966). The outer lip is white to chocolate-brown in colour. All the specimens that we investigated correspond to this morphotype although two individuals had only one vague band on the lower half of the body whorl.
In Rhynchotrochus williamsi the penis is much shorter than the vas deferens and is shorter or of the same length as the epiphallus. The penis consists of a very thick, muscular proximal part and a more slender distal part that is somewhat shorter than the proximal part. The oviduct is approximately half as long as the vagina. Rhynchotrochus williamsi has a longer epiphallus and vas deferens compared to R. taylorianus although individuals of R. taylorianus from Blup Blup and Madang also have a long vas deferens. The shell of R. williamsi is globose and white with a white to light pink outer lip. The spiral bands show only as translucent areas. The protoconch is usually a dark brown (Clench & Turner, 1966). The four individuals from Laing Island corresponded with this morphotype.
The reproductive anatomy of R. taylorianus is very similar to that of R. williamsi but in the former, the epiphallus is shorter than the vas deferens and the LO is approximately two-thirds that of the vagina. Especially the LVd, epiphallus and bursa copulatrix show considerable intraspecific variation. Clench & Turner (1966) reported R. taylorianus to be extremely variable for shell size, shell colour and shell banding pattern: the shell is subdepressed and trochiform, generally yellowish with pinkish purple on the base and on the upper whorls; some shells have spiral bands both above and below the periphery that consist of purple or brown flecks; the outer lip is dark purplish-black and the interior of the shell is white to dark purple. Not all species have the purple-brown colour pattern, but instead have pinkish or white older whorls and a yellow body whorl. The protoconch is coloured either light or dark. Remarkably, only the four individuals from Madang had the typical dark purplish-black outer lip, all the other individuals had a white outer lip. Individuals from four localities were very similar in shell colour and banding pattern. Except for the number of shell bands, individuals from each locality were very similar in colour.
Most of the Rhynchotrochus species are described on a few specimens and are discriminated on shell shape, shell colour and shell banding pattern only. Our study also shows that the three Rhynchotrochus species can be reliably discriminated using the colour and banding pattern of the shell alone. Rhynchotrochus species are best regarded as morphospecies since no other species concept, such as the widely and commonly applied biological species concept, has ever been tested in this genus. Hence, unless we know the genetics of the variation in shell colour and banding pattern these characters should be carefully applied as species markers. For instance, it is unknown whether the striking difference in colour among individuals from several of the localities of R. taylorianus represents intraspecific variation through fixation of different alleles coding for shell colour and banding pattern, or whether these differences are the result of differentiation resulting from reproductive isolation. Further, we added descriptions of the reproductive organs of three species, but for the majority of species (i.e. 14 out of 20) such descriptions are lacking. Finally, additional potentially valuable taxonomic markers should be evaluated such as the radula, the spermatophore and the internal structure of the penis.
Most anomalies of the reproductive organs in pulmonate land snails relate to the male parts. Abnormal male reproductive organs have been reported in Helix pomatia Linné, 1758 (Paravicini, 1898; Pėgot, 1900; Ashworth, 1907; Lattmann, 1967), Cernuella virgata (Da Costa, 1778) (Hausdorf, 1989) and Mastus cf. cretensis (Pfeiffer, 1846) (Maassen, 2000). The female organs were always normally developed. In H. pomatia and C. virgata, abnormal individuals may have three sets of male copulatory organs. One set is always complete with a penis, an epiphallus, an appendix, flagellum and retractor muscle whereas one or more of these structures are lacking in both other sets. The situation in R. taylorianus is most similar to that of M. cf. cretensis. In both species, two penes and epiphalli are connected with a single, short vas deferens that does not make contact with the spermoviduct. Yet, in M. cf. cretensis the proximal parts of both penes are interconnected and do not make contact with the genital atrium, whereas in R. taylorianus both penes insert at the genital atrium, each at one side. Successful copulation as a male is impossible in M. cf. cretensis and R. taylorianus, but since the female organs are normally developed and connected with the genital atrium, copulation as a female should be possible, unless simultaneous reciprocal intromission of penes is necessary for successful copulation. Copulation as male and female is prevented in C. virgata where only a blind-ending penis inserts at the genital atrium (Hausdorf, 1989). In H. pomatia, successful copulation as male and female, in theory, is possible but could be accompanied by practical difficulties (Pėgot, 1900).
ACKNOWLEDGEMENTS
We wish to thank Harry Van Paesschen for the artwork and an anonymous referee for improving the manuscript. K.J. is a postdoctoral fellow at the Fund for Scientific Research – Flanders (F.W.O.).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}